몫:



펜 토즈 단계 및 관련 질병의 경로
그 오색의 길 또한 착신 모노 포스페이트 탄당라고도 포스페이트, 그 최종 제품 뉴클레오티드와 같은 DNA, RNA, ATP, NADH, FAD 및 조효소 A 등의 핵산의 합성 경로에 필요한 리보스 기본적인 대사 경로이다.
그것은 또한 다양한 효소 반응에 사용되는 NADPH (nicotinamide adenine dinucleotide phosphate)를 생산합니다. 이 경로는 매우 동적이며 세포의 순간적 요구에 따라 제품을 적용 할 수 있습니다..
ATP (adenosine triphosphate)는 가수 분해가 광범위한 생화학 반응에 결합 될 수 있기 때문에 세포의 "에너지 통화"로 간주됩니다.
마찬가지로, 제 NADPH를 포함한 환원성 지방산의 합성 콜레스테롤 합성, 신경 전달 물질 합성, 광합성 해독 반응 에너지 통화 필요.
NADPH와 NADH는 구조가 유사하지만 생화학 반응에서 서로 바꾸어 사용할 수는 없습니다. NADPH는 환원성 생합성을위한 특정 대사 산물의 산화에서 자유 에너지의 사용에 참여한다.
반대로, NADH는 ATP를 합성 대사 산화 자유 에너지의 사용에 관여.
색인
- 1 역사 및 위치
- 2 함수
- 3 단계
- 3.1 산화 상태
- 3.2 비 산화 상
- 4 관련 질병
- 5 참고
연혁과 위치
이 루트의 존재의 징후는 1930 년 Otto Warburg 수사관 덕분에 시작되었습니다. NADP의 발견은 그에게 귀속됩니다+.
어떤 관찰은 그 경로의 발견, 특히 불소 이온과 같은 해당 과정의 억제제가있을 때 호흡의 지속을 허용했다.
1950 년 과학자 Frank Dickens, Bernard Horecker, Fritz Lipmann 및 Efraim Racker는 오탄당 인산염 경로.
유방 땀샘, 지방 조직 및 신장과 같은 콜레스테롤과 지방산의 합성에 관여하는 조직은 높은 농도의 오탄당 인산 효소.
간은 또한이 경로에 중요한 조직입니다.이 조직에서 포도당 산화의 약 30 %는 오탄당 인산염 경로의 효소 덕분에 발생합니다.
기능들
오탄당 인산염 경로는 세포에서 탄소의 항상성을 유지하는 역할을합니다. 마찬가지로, 경로는 아미노산 (펩타이드와 단백질의 구조 블록)의 합성에 관여하는 뉴클레오티드와 분자의 전구체를 합성하고,.
그것은 효소 반응을 감소시키는 주요 원천입니다. 또한 단백 동화 반응과 산화 스트레스에 대한 방어 과정에 필요한 분자를 제공합니다. 트랙의 마지막 단계는 스트레스 상황에서 산화 환원 공정에서 중요합니다..
단계
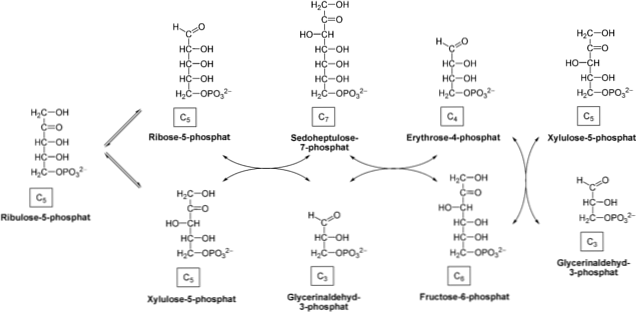
오탄당 인산염 경로는 세포질 세포질에서 두 단계로 구성된다 : 포도당 -6- 인산염이 리보스 -5- 인산염으로 산화되어 NADPH를 생성하는 산화 적 산화 환원법; 및 3, 4, 5, 6 및 7 개의 탄소의 당의 상호 전환을 수반하는 비 산화.
이 경로는 칼빈 (Calvin) 순환 및 해당 과정에 대한 대안 인 Entner-Doudoroff 경로와 공유되는 반응을 나타냅니다.
산화 단계
산화 적 탈수 소화상은이 반응은 NADP에 대해 높은 특이성을 가진 효소 글루코스 -6- 포스페이트 탈수소 효소에 의해 촉매된다 1. 탄소 분자의 글루코오스 -6- 포스페이트로 시작+.
이 반응의 생성물은 6- 포스 포노 글루 코노 -δ- 락톤이다. 이어서,이 생성물을 효소 락 타나 제로 가수 분해시켜 6- 포스 포 글루코 네이트를 얻는다. 후자의 화합물은 효소 6- 포스 포 글루코 네이트 탈수소 효소에 의해 취해지고 리브 로스 5- 인산.
phosphopentose isomerase 효소는 ribulose 5-phosphate의 이성체 화에 의한 ribose 5-phosphate의 합성을 포함하는 산화 단계의 마지막 단계를 촉매한다.
이 일련의 반응은 글루코오스 6 인산 분자 1 분자 당 2 분자의 NADPH와 1 분자의 리보오스 5 인산염을 생성하여이 효소 경로로 들어간다..
일부 세포에서는 NADPH 요구량이 리보오스 5- 인산보다 높다. 따라서 transketolase transaldolase 및 효소가 리보스 -5- 인산을 비산 화성 및 위상 선도, 글리 세르 알데히드 -3- 포스페이트와 프룩 토스 -6- 포스페이트로 변환한다. 이 마지막 두 가지 화합물은 당분 해 경로로 들어갈 수 있습니다..

비산 화성 상
상은 효소 펜 토스 -5- 인산 에피 머라 아제에 의해 촉매 화 된 에피 머화 반응으로 시작한다. Ribulose-5- 인산염은이 효소에 의해 취해지고 xylulose-5-phosphate.
생성물은 리보스 -5- 포스페이트를하기 크실 로스 5- 인산의 통과를 촉진 조효소 티아민 피로 포스페이트 (TPP)와 함께 작용 transketolase 효소에 의해 취해진 다. 케도 오스를 알도 오스로 이동 시키면, 글리 세르 알데히드 -3- 인산 및 세도 헵 룰로오스 -7- 인산이 생성된다.
이어서 transaldolase 효소는 C3 분자는 4 탄당을 생산 글리 세르 알데히드 -3- 포스페이트 (에리트 -4- 포스페이트) 내지 7 인산 여섯 카본 (프 룩토 오스 -6- sedoheptulose 전송 - 인산염). 이 제품은 당분 해 경로를 먹일 수 있습니다..
효소는 C2에 크실 로스를 에리트 -4- 포스페이트를하려면 -6- 포스페이트를 과당 글리 세르 알데히드 -3- 포스페이트 결과로 이어지는 5 인산을 전송할 다시 transcetosala 작용한다. 이전 단계에서와 마찬가지로 이들 제품은 해당 과정에 들어갈 수 있습니다.
이 두 번째 단계는 NADPH를 생성하는 경로와 ATP와 NADH를 합성하는 경로를 연결합니다. 또한, 프럭 토스 -6- 포스페이트 및 글리세리드 알데히드 -3- 포스페이트는 글루코 네오 신.
관련 질병
다른 병리학은이 신경근 질환과 다른 유형의 암 사이의 오탄당 인산염의 경로와 관련이있다.
대부분의 임상 연구는 글루코스 -6- 포스페이트 탈수소 효소의 활성을 정량화하는데 초점을 맞추고 있는데, 이는 통로 조절에 관여하는 주요 효소이기 때문이다.
빈혈에 걸리기 쉬운 개인에 속한 혈액 세포에서 글루코오스 -6- 인산 탈수소 효소의 효소 활성이 낮습니다. 대조적으로, 후두의 암종과 관련된 세포주는 높은 효소 활성을 나타낸다.
NADPH는 산화 스트레스에 관여하는 활성 산소 종에 대한 보호에 중요한 펩타이드 분자 인 글루타티온 생산에 관여한다.
다른 유형의 암은 오탄당 경로의 활성화로 이어지고 전이, 혈관 신생 및 화학 요법 및 방사선 치료에 대한 반응과 관련이 있습니다.
다른 한편, 만성 육아 종성 질환은 NADPH 생산에 결함이있을 때 발생합니다.
참고 문헌
- Berg, J.M., Tymoczko, J.L., Stryer, L (2002). 생화학. WH 프리먼
- Konagaya, M., Konagaya, Y., Horikawa, H., & Iida, M. (1990). 신경근 질환에서 Pentose phosphate pathway - 근육 포도당 6- 인산 탈수소 효소 활성 및 RNA 함량 평가. Rinsho shinkeigak. 임상 신경학, 30(10), 1078-1083.
- Kowalik, M.A., Columbano, A., & Perra, A. (2017). 간세포 암에서의 오탄당 인산염 경로의 새로운 역할. 종양학의 최전선, 7, 87.
- Patra, K. C., & Hay, N. (2014). 오탄당 인산염 경로와 암. 생화학 분야의 동향, 39 세(8), 347-354.
- Stincone, A., Prigione, A., Cramer, T., Wamelink, M., Campbell, K., Cheung, E., & Keller, M. A. (2015). 신진 대사의 재현 : 오탄당 인산염 경로의 생화학 및 생리학. 생물학 리뷰, 90(3), 927-963.
- Voet, D., & Voet, J. G. (2013). 생화학. Artmed Publisher.