몫:



Anticodon 설명, 기능 및 코돈과의 차이점
A 안티코돈 전사 RNA 분자의 그 함수의 메신저 RNA 분자 내에 존재하는 세 개의 다른 뉴클레오티드 서열을 인식한다 (tRNA에)에 존재하는 세 개의 뉴클레오티드 서열 (mRNA의)입니다.
코돈과 안티코돈 사이의 이러한 인식은 역 평행하다. 즉, 하나는 5 '→ 3'방향에 위치하고 다른 하나는 3 '→ 5'방향에 있습니다. 세 개의 뉴클레오타이드 (세 쌍)의 염기 서열 사이의 인식은 번역 과정에서 기본적입니다. 즉, 리보솜에있는 단백질의 합성에서.
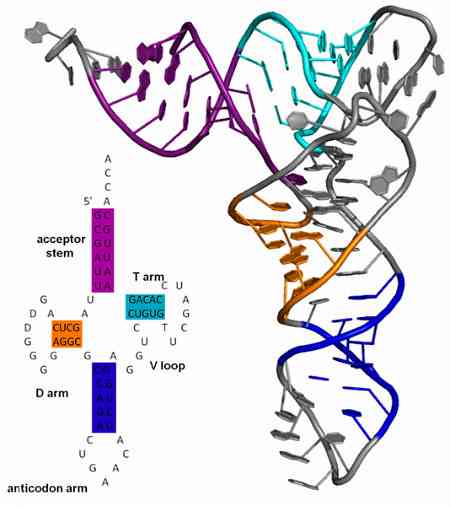
따라서 번역하는 동안 전령 RNA 분자는 전달 RNA의 안티코돈 (anticodons)에 의해 코돈의 인식을 통해 "판독"됩니다. 이 분자들은 특정 아미노산을 리보솜에서 형성되는 단백질 분자로 옮기기 때문에 그렇게 불립니다.
20 개의 아미노산이 있으며 각각은 특정 삼중 항으로 인코딩됩니다. 그러나 일부 아미노산은 하나 이상의 삼중 항으로 인코딩됩니다.
또한 일부 코돈은 아미노산이 붙어 있지 않은 전달 RNA 분자의 안티코돈 (anticodons)에 의해 인식된다. 이들은 소위 정지 코돈이다..
색인
- 1 설명
- 2 함수
- 안티코돈과 코돈의 차이점
- 4 롤링의 가설
- 4.1 RNA와 아미노산
- 5 참고
설명
아데닌 (A), 구아닌 (G), 우라실 (U) 또는 시토신 (C) 세 개의 뉴클레오티드의 조합되도록 일 : 안티코돈은 다음 질소 염기를 포함 할 수있는 세 개의 뉴클레오티드 서열로 구성 그것은 코드로 작동.
안티코돈은 항상 RNA 분자에서 발견되며 항상 3 '-> 5'방향으로 놓여 있습니다. 이러한 tRNA의 구조는 네 개의 루프 (또는 루프)로 세분화 된 방식으로 클로버와 유사합니다. 루프 중 하나에서 안티코돈이 있습니다..
안티코돈은 전령 RNA의 코돈을 인식하고, 결과적으로 모든 살아있는 세포에서 단백질 합성 과정에 필수적이다.
기능들
안티코돈의 주된 기능은 메신저 RNA 분자에서 코돈을 형성하는 삼중 항의 특정 인식이다. 이 코돈은 DNA 분자에서 복사되어 단백질의 아미노산 순서를 지시하는 지침입니다.
전사 (메신저 RNA 사본의 합성)가 5 '→ 3'방향에서 발생하기 때문에, 전달 RNA의 코돈은 이러한 방향을 갖는다. 따라서, 전달 RNA 분자에 존재하는 안티코돈은 반대 방향 인 3 '→ 5'.
이 조합은 상보성 때문입니다. 예를 들어, 한 코돈이 5'-AGG-3 '이면 안티코돈은 3'-UCC-5'입니다. 코돈과 안티코돈 간의 이러한 특이한 상호 작용은 메신저 RNA의 뉴클레오타이드 서열이 단백질 내의 아미노산 서열을 암호화 할 수있게하는 중요한 단계입니다..
안티코돈과 코돈의 차이점
- 안티코돈은 tRNA에서 mRNA의 코돈과 상보적인 3 염기쌍이다. 단백질 생산 과정에서 tRNA가 정확한 아미노산을 전달할 수 있습니다. 대조적으로, 코돈은 단백질 합성에서 특정 아미노산을 코딩하는 DNA 또는 mRNA에서 trinucleotides의 단위입니다.
- 안티코돈은 mRNA의 뉴클레오티드 서열과 단백질의 아미노산 서열 사이의 연결이다. 반대로, 코돈은 DNA가있는 핵에서 단백질 합성이 일어나는 리보솜으로 유전 정보를 전달합니다..
- 안티코돈은 DNA와 mRNA 분자에 위치한 코돈과 달리 tRNA 분자의 안티 코돈 (antiicodon)에서 발견된다.
- 안티코돈은 각각의 코돈에 상보 적이다. 대조적으로, mRNA의 코돈은 DNA 내의 특정 유전자의 뉴클레오티드의 삼중 항에 상보 적이다.
- tRNA는 안티코돈 (anticodon)을 포함하고 있습니다. 대조적으로, mRNA는 다수의 코돈.
롤링 가설
가설 코돈 메신저 RNA의 세 번째 뉴클레오티드 및 전사 된 RNA 안티코돈의 첫번째 뉴클레오티드의 접합 압연 제안하고 다른 두 뉴클레오타이드 삼중 접합의보다 구체적인.
Crick은이 현상을 각 코돈의 세 번째 위치에서 "흔들리는"것으로 묘사했습니다. 그 위치에서 노동 조합이 평범한 것보다 덜 엄격해질 수 있습니다. 그것은 또한 흔들림 (wobbling) 또는 탐 볼레 (tamboleo)라고도합니다..
이 크릭 워블 가설은 주어진 tRNA의 안티코돈이 2 개 또는 3 개의 상이한 mRNA 코돈과 어떻게 짝을 이룰 수 있는지를 설명한다.
크릭 평소보다 덜 엄격한 (의 tRNA 안티코돈의베이스 (59)와베이스 (39)의 mRNA의 코돈 사이) 염기쌍은이 사이트 "워블"또는 감소 된 친 화성 특정 허용 될 때, 제안.
결과적으로, 단일 tRNA는 주어진 아미노산을 특정하는 두 개 또는 세 개의 관련된 코돈을 종종 인식합니다.
일반적으로, tRNA 안티코돈의 염기와 mRNA 코돈 사이의 수소 결합은 코돈의 처음 두 염기에 대해서만 엄격한 염기쌍 규칙을 따른다. 그러나,이 효과는 모든 mRNA 코돈의 모든 세 번째 위치에서 일어나지는 않는다.
RNA와 아미노산
워블 가설에 기초하여 완전한 변성을 나타내는 코돈을 갖는 각 아미노산에 대해 적어도 2 개의 전달 RNA의 존재가 예측되었는데, 이는 사실로 입증되었다.
이 가설은 또한 6 개의 모든 세린 코돈에 대해 3 개의 전달 RNA의 출현을 예측했다. 실제로 세린에 대한 3 개의 tRNA가 특성 분석되었습니다.
- 세린 1 (anticodon AGG)에 대한 TRNA는 코돈 UCU 및 UCC에 결합한다.
- 세린 2 (anticodon AGU)에 대한 TRNA는 코돈 UCA 및 UCG에 결합한다.
- 세린 3 (anticodon UCG)에 대한 TRNA는 코돈 AGU 및 AGC에 결합한다.
이러한 특이성은 정제 된 아미노 아실 -tRNA 트리 뉴클레오타이드가 시험 관내에서 리보솜으로의 자극 된 결합에 의해 확인되었다.
마지막으로, 몇몇 transfer RNA는 hypoxanthine purine으로 만든 이노신 염기를 가지고있다. 이노 신은 아데노신의 전사 후 변형에 의해 생성된다.
크릭 스 태거 가설 이노신 코돈에서 우라실, 시토신 또는 아데닌과 일치하는 것 인 안티코돈 (요동 위치)의 5 '말단에 존재하는 경우, 예측.
실제로, 안티코돈의 5 '위치에있는 이노신 (I)을 포함하는 정제 된 알라 닐 -tRNA는 GCU, GCC 또는 GCA의 트리 뉴클레오타이드로 활성화 된 리보솜에 결합한다.
안티코돈의 5 '위치에서 이노신으로 정제 된 다른 tRNA에서도 동일한 결과가 얻어졌다. 그러므로 Crick의 워블 가설은 유전 암호가 주어진 tRNA와 코돈 사이의 관계를 매우 잘 설명한다..
참고 문헌
- Brooker, R. (2012). 유전학의 개념 (1 판). McGraw-Hill Companies, Inc.
- Brown, T. (2006). 게놈 3 (3rd). 갈랜드 과학.
- Griffiths, A., Wessler, S., Carroll, S. & Doebley, J. (2015). 유전 분석 입문 (11 판). W.H. 프리먼
- 루이스, R. (2015). 인간 유전학 : 개념과 응용(11 판). McGraw-Hill 교육.
- Snustad, D. & Simmons, M. (2011). 유전학의 원리(6 판). 존 와일리와 선즈.